All true love must dieSo it goes.
Alter at the best
Into some lesser thing.
Prove that I lie.
Such body lovers have,
Such exacting breath,
That they touch or sigh.
Every touch they give,
Love is nearer death.
Prove that I lie.
Tuesday, 26 January 2010
Cleansing by writing
[deleted]
Tuesday, 22 September 2009
One Night at Victoria Beach
I typed it up for another forum, but I thought it would be a good idea to archive here as well, I guess. This was one of my childhood favourites:
[B]One Night at Victoria Beach[/B]
The wind comes rushing from the sea,
the waves curling like mambas strike
the sands and recoiling hiss in rage
washing the Aladuras' feet pressing hard
on the sand and with eyes fixed hard
on what only hearts can see, they shouting
pray, the Aladuras pray; and coming
from booths behind, compelling highlife
forces ears; and car lights startle pairs
arm in arm passing washer-words back
and forth like haggling sellers and buyers -
Still they pray, the Aladuras pray
with hands pressed against their hearts
and their white robes pressed against
their bodies by the wind; and drinking
palm-wine and beer, the people boast
at bars at the beach. Still they pray.
They pray, the Aladuras pray
to what only hearts can see while dead
fishermen long dead with bones rolling
nibbled clean by nibbling fishes, follow
four dead cowries shining like stars
into deep sea where fishes sit in judgement;
and living fishermen in dark huts
sit around dim lights with Babalawo
throwing their souls in four cowries
on sand, trying to see tomorrow.
Still they pray, the Aladuras pray
to what only hearts can see behind
the curling waves and the sea, the stars
and the subduing unanimity of the sky
and their white bones beneath the sand
And standing dead on dead sands,
I felt my knees touch living sands-
but the rushing wind killed the budding words.
- Gabriel Okara
[B]One Night at Victoria Beach[/B]
The wind comes rushing from the sea,
the waves curling like mambas strike
the sands and recoiling hiss in rage
washing the Aladuras' feet pressing hard
on the sand and with eyes fixed hard
on what only hearts can see, they shouting
pray, the Aladuras pray; and coming
from booths behind, compelling highlife
forces ears; and car lights startle pairs
arm in arm passing washer-words back
and forth like haggling sellers and buyers -
Still they pray, the Aladuras pray
with hands pressed against their hearts
and their white robes pressed against
their bodies by the wind; and drinking
palm-wine and beer, the people boast
at bars at the beach. Still they pray.
They pray, the Aladuras pray
to what only hearts can see while dead
fishermen long dead with bones rolling
nibbled clean by nibbling fishes, follow
four dead cowries shining like stars
into deep sea where fishes sit in judgement;
and living fishermen in dark huts
sit around dim lights with Babalawo
throwing their souls in four cowries
on sand, trying to see tomorrow.
Still they pray, the Aladuras pray
to what only hearts can see behind
the curling waves and the sea, the stars
and the subduing unanimity of the sky
and their white bones beneath the sand
And standing dead on dead sands,
I felt my knees touch living sands-
but the rushing wind killed the budding words.
- Gabriel Okara
Indulgence
The one where I take time out to feel sorry for myself
I know I've fucked up. In fact, if one were to be perfectly honest, I am a fuck-up. She didn't have to say it - none of them did; the silences and pauses said it all, clearly - plainly. There's probably little that can be politely said to the loser that refuses to get back up from the ground. Therein lies the problem: that I have chosen to remain on the ground for so long, content with scraps, and constantly scared and ashamed even of my own shadow. She was right to dismiss me and everything I said. I could find no argument. And even though I could have given excuses, I could offer no argument. Reasons and caveats piled up in mind, but I could offer up no arguments. She was right and I acceded. She wants to part and I saw in it her thinking that it might be the best way for her to keep her sanity. So it goes. No excuses.
Having said all that, the rot stops now. I will arise out of my rut, go farther than I have gone before, and say "I wish you had believed in me despite all that had happened. I wish you could have conjured up just a little faith when I asked for... a little rope." Those declarations will not come from a place of malice, just from a realisation.
It is time to wake up. It's time I rediscovered my confidence. No excuses, just existence.
I know I've fucked up. In fact, if one were to be perfectly honest, I am a fuck-up. She didn't have to say it - none of them did; the silences and pauses said it all, clearly - plainly. There's probably little that can be politely said to the loser that refuses to get back up from the ground. Therein lies the problem: that I have chosen to remain on the ground for so long, content with scraps, and constantly scared and ashamed even of my own shadow. She was right to dismiss me and everything I said. I could find no argument. And even though I could have given excuses, I could offer no argument. Reasons and caveats piled up in mind, but I could offer up no arguments. She was right and I acceded. She wants to part and I saw in it her thinking that it might be the best way for her to keep her sanity. So it goes. No excuses.
Having said all that, the rot stops now. I will arise out of my rut, go farther than I have gone before, and say "I wish you had believed in me despite all that had happened. I wish you could have conjured up just a little faith when I asked for... a little rope." Those declarations will not come from a place of malice, just from a realisation.
It is time to wake up. It's time I rediscovered my confidence. No excuses, just existence.
Sunday, 20 September 2009
One Step Forward, Two Steps Backwards.
The teachings are sheer fantasy, unsubstantiated by any credible evidence: ancient Egyptians mastered flight with gliders, which they used for both recreation and travel. They invented electric batteries and mastered electroplating, discovered the principles of quantum mechanics and anticipated Darwin's theories of evolution. Furthermore, all Egyptians were black, and their abundance of the dark skin pigment, melanin, not only made them more humane and superior to lighter-skinned people in body and mind but also provided such paranormal powers as ESP and psychokinesis.
Incredible as it may seem, these fallacies are being included in public school multicultural courses in a growing number of U.S. cities and espoused in black-studies departments on some college campuses.
More: http://www.time.com/time/magazine/article/0,9171,980453-1,00.html
Afrocentric studies in American schools are full of pseudoscience and total bollocks? Dolts!
Wednesday, 22 July 2009
For KAG, wherever he or she may be.
A song of David.
1. The lost wind finds a way across the plains, its breath settling for paths unknown, unconquered, previously hidden from sight. 2. In the midst of all the razed, mighty Iroko stood upright, unshaken by Zephyr's call. 3. In the face of catastrophe, mighty Iroko, unfazed and calm, shed some weight but held arms high: ready for battle. 4. Let us salute the movement that brings music. 5. Let us salute the standing that brings difference. 6. It is true all trees fall. It is true all trees are temporarily vanquished. All those and more are true with time. 7. Yet, there is no perishing; all change into something anew. 8. See the rise of new sprouts! 9. See mighty Iroko with new faces all bright standing again in the sight of new land 10. Let us praise the wind. Let us joyfully proclaim its power 11. At the end, though, we shall not forget mighty Iroko, difference and arising.
1. The lost wind finds a way across the plains, its breath settling for paths unknown, unconquered, previously hidden from sight. 2. In the midst of all the razed, mighty Iroko stood upright, unshaken by Zephyr's call. 3. In the face of catastrophe, mighty Iroko, unfazed and calm, shed some weight but held arms high: ready for battle. 4. Let us salute the movement that brings music. 5. Let us salute the standing that brings difference. 6. It is true all trees fall. It is true all trees are temporarily vanquished. All those and more are true with time. 7. Yet, there is no perishing; all change into something anew. 8. See the rise of new sprouts! 9. See mighty Iroko with new faces all bright standing again in the sight of new land 10. Let us praise the wind. Let us joyfully proclaim its power 11. At the end, though, we shall not forget mighty Iroko, difference and arising.
Tuesday, 2 June 2009
Winace's excellent shared ERVs primer part 2
3. Attempted creationist rebuttals and their drawbacks
Hypotheses proposed by creationists to account for this are woefully inadequate, although there are several worth analyzing.
3.1: Independent Insertion
This one entails independent insertion by the same virus affecting different species. Creationist Ashby Camp, writing at TrueOrigins, quoted a scientific article referring to "insertion hotspots" that were the target of viral integration much more often than other spots in the genome. Although this is superficially a valid criticism to be hurled at the ERV argument, it quickly fails.
The very "hottest" spot Camp could dig up in the scientific literature was one that was 280 times more likely to be involved in a viral integration than we could expect from chance alone. It could be pointed out that this was a virus that doesn't exist in nature, but was specifically designed to facilitate gene therapy by targeting a specific part of the genome and replacing a crippled gene with a functional copy.
But let's ignore that for now, and assume Camp did not misrepresent relevant research. Just how large is this in the big picture? The human genome is 3 billion or so nucleotides long (Source). The chance, then, of randomly inserting into the same section of the genome is 1 in 3 billion.
Now let's assume there are hot spots an unprecedented 1,000 times more likely to be attacked by a virus than the unique, genetically engineered one Mr. Camp was able to dig up. Divide 3 billion by 280,000, and you find the chance of an independent insertion is one in around 10714; this is an unlikely enough figure to be negligible, and it was derived from estimates orders of magnitude more liberal than the data would allow!
Additionally, this doesn't address any of the other factors involved. It doesn't explain why the same viral fingerprint would be left behind, how it would affect the one germ line cell out of millions that advances to zygote status, how that single individual would enjoy reproductive success sufficient to establish his ERVs in the population, etc.
For the reasons listed above, it's astronomically improbable. Not only would the same exact retroviral insertion have to occur independently at the same exact locus of a uniquely lucky sperm or egg cell that survived to adulthood and achieved enough success to establish the insertion in their respective populations, but this impossible set of coincidences would have to occur for 3, 4, even a dozen species at the same time, and for every single ERV shared between them to boot!
As you can see, the improbabilities keep stacking and increasing by additional orders of magnitude as more factors are introduced.
The additional fact that ERVs, when analyzed, yield evolutionary patterns that reinforce the phylogenies derived from morphological, fossil and other evidence rules out the independent insertion hypothesis completely.
If independent insertion was somehow the explanation for shared viral insertions, we would expect to see numerous, extensive groups of ERVs common to chickens and humans, pigs and humans, and other creatures that share many of the same pathogens; ERVs that weren't present, on the other hand, in geographically isolated creatures closer to us in morphology and genetics, such as the apes.
3.2: ERVs are not remnants of ancient viral infection
Another attempted explanation is that the ERVs found in our genome are actually original, designed artifacts instead of viral infection fingerprints.
To be frank, this hypothesis, even on the surface, appears as ludicrous as assertions that dinosaur bones don't actually come from dinosaurs but were intentionally placed there. Science generally doesn't deal with Omphalos hypotheses for good reason, because there is no reasonable way to verify or falsify them; they throw explanatory and predictive power, much less parsimony, entirely out the window.
People who assert this for ERVs have the burden of proof in demonstrating where shared ancestry ends and intentional design begins. Are the various breeds of domestic cat different, independently created organisms? (They share ERVs that have been used to reconstruct their phylogenies). Are the various felines in general related, or seperately created? (Lions, panthers, tigers and domestic cats share ERVs).
In other words, where do the divinely faked ERVs end and the real ones, which we can observe being incorporated into the genome in vitro, begin? ERV insertion is a well-documented event, leaving very specific and unlikely patterns; no other process except viral infection has been documented that can create them.
Why, then, should any reasonably intelligent person consider accepting rationalizations that are unfalsifiable, unevidenced, predict no unique observations the mainstream explanation doesn't and even require numerous additional assumptions that the obvious answer doesn't?
Nevertheless, let us examine two sub-hypotheses that fall under this main heading.
3.2.1: ERVs are an important part of the immune system
Based on the fact that a few ERVs have apparently been shown to lessen the chances of some types of viral infection, some creationists have asserted that ERVs are actually an anti-viral vector native to the genome, with their similarities to actual retroviruses being coincidental and not the product of intentional deception. But this explanation also fails when all the data is considered.
In the first place, their anti-viral effects are no larger than those of some free-living viruses, so this is not an argument against their being parasitic in the past. Human herpesvirus 6 has recently been shown to suppress HIV (source), for example. At most, it would provide a miniscule selection advantage for the individual carrying a particular ERV, which could help it piggyback across the population and help slightly improve the odds for one step of the process (out of many) occuring independently.
A possible explanation is that the endogenous retrovirus, which is only marginally deleterious, uses up the spaces on the cellular exterior HIV would normally dock with. This would be a textbook example of an evolutionary, jury-rigged defense mechanism, equivalent to hiring polite burglars to slowly work on your doors and windows so the violent ones couldn't get in.
Additionally, virtually all ERVs have no such immunity-boosting function; indeed, many more are actually implicated as the cause of some tumors (source); just like certain free-living viral infections including Epstein-Barr, I might add.
Moreover, you again have the problem that ERVs are distributed across the genomes of species in a pattern that mimicks common descent, not the expected pattern of functionally-oriented resistance to shared viral pathogens. I can safely predict that chickens and humans won't have any common anti-viral ERV that apes won't, despite the fact they share many of the same infections apes generally aren't afflicted by.
3.2.2: Viruses evolved from ERVs, not the other way around
Some have asserted that ERVs were actually designed elements that exogenous retroviruses came from, perhaps after the Fall. This is advocated by creationist Dr. Ian Macreadie at this AnswersInGenesis article, who states "I actually don’t believe God created viruses as separate entities, I believe they were a part of the DNA in cells".
A few things are to be noted. As this webpage demonstrates, this is a modern reinvention of an ancient, proto-scientific view aimed at explaining the existence of regular, plain vanilla, non-molecular fossils:
"... they were owing to the actions of so-called 'plastic forces' or 'formative virtues' in the Earth... 'Plastic forces' were believed to be unspecified forces within the Earth that were continually striving to produce organic bodies. The notion that fossils were formed by these 'plastic forces', first put forward by the Arab scholar Avicenna (980-1037 AD), was an outgrowth of the Aristotelian idea of spontaneous generation. Fossils were individuals that had 'failed' at spontaneous generation, or else the 'vital essence' of living things had penetrated rocks and formed objects there which resembled living things."
We now know much more about the world, and recognize that fossils are the remnants of organisms, not their ancestors. Can we similarly argue that ERVs can't be the parents of ordinary viruses?
As it turns out, the answer is a resounding "Yes!" This creationist hypothesis is unambiguously falsified by looking at the data.
Many ERVs are merely fragments of code, which nevertheless include readily vestiges of identifiable viral surface coat proteins, which are quite obvious and expected for an exogenous retrovirus that needs to float around independently, but utterly baffling for one that resides within a cell.
In other words, they lack key components that allow real viruses to reproduce (as a side note, this is often, in the first place, a relic of a botched infection, one of the things that lets the cell survive in the first place).
If a sentence out of Shakespeare is inserted into an essay, one can ascertain the direction of literary borrowing from comparison of the works alone, even without the knowledge that the former wrote earlier. This is the same with ERVs and other out-of-place genetic fragments. They simply look like fragments out of the whole work, so to speak, deposited there by free-living ancestors, not the opposite.
To expand even further and bring in the heavy artillery, many viruses have complex, multipart mechanisms for subverting host immune defenses and injecting their DNA thru the cell membrane that their fragmentary, cell-bound counterparts lack.
The bacteriophage T4 virus has a head, tail, baseplate and a dozen tail fibers. The baseplate serves as a "nerve center" of the virus. When the tail fibers attach to E. coli, the baseplate transmits a message to the tail, which contracts like a muscle, bringing the internal pinlike tube in contact with the outer membrane of the E. coli cell. As the tube punctures the outer and inner membranes, the virus' DNA is injected into the host cell. (Source)
Needless to say, asserting that such things could evolve from mere cellular genome fragments is a stretch, especially so if one simultaneously denies the ability of evolutionary mechanisms to generate such staggering complexity.
Finally, a customary red herring involved in any discussion of molecular evidence for evolution is cries of "but they have a FUNCTION!". While that may well be true, function (or lack of it) is generally not the criterion by which things are considered evidence for evolution, as demonstrated by the case I made above.
4. Conclusion
In summary, the facts are that:
(A) A retrovirus infects a cell and, depending on how virulent it is, can kill it. Occasionally, fragments of the virus remain but the cell survives.
(B) No two viral insertions are exactly alike; a botched integration can leave behind a tiny fragment of its DNA, large stretch of the genome, selected portions, and other variants.
(C) Viral fragments insert at fairly random locations, and have literally millions of them to choose from.
(D) Infection of a germ line cell is quite rare.
(E) That particular germ line cell with the unique viral fingerprint can, sometimes, be the lucky egg or sperm cell that gets fertilized.
(F) The individual's genes, including the ERV, can get established in the entire population of a species. This can occur through random genetic drift if it's neutral or deleterious; natural selection, on the other hand, would work to establish the very few advantageous retroviral elements in our genome.
The combined odds of each of these events happening in two independently originated species are a statistical non-event. In sheer unlikelihood, they would likely dwarf even the incorrectly calculated anti-abiogenesis calculations some creationists are fond of. And the odds become even less remote, by additional orders of magnitude, as more and more species are considered.
In this post, we've seen that ERVs are, indeed, remnants of ancient viral infection. We've also seen that no explanation except inheritance from a common ancestor can reasonably account for the occurence of the same viral insertion in two different organisms or species. This, in turn, leads to the inescapable, iron-clad, demonstrated-beyond-a-reasonable-doubt conclusion:
Independent origin of ERVs (and hence, independent origin of species, a.k.a. creationism) is scientifically falsified.
P.S. Before the resident creationists link to http://www.trueorigin.org/theobald1e.asp , please attempt to dialogue with the refutation I layed out above.

Sauce: http://www.christianforums.com/t96639/#post1828403
Winace's excellent shared ERVs primer part 1
The last time I mentioned ervs, I should have posted the late, great Winace's excellent primer on ERVs. Let's make things right. I'm copy/pasting from the one that he originally posted on the old Christian Forums. It's an oldie, but a goodie.
ERVs, a startling line of evidence from genetics (not fossils) provides additional, overwhelming support for human/chimp common ancestry, one that establishes it beyond a shadow of a doubt.
Much like stellar parallax could be considered 'proof' of heliocentricism, so too do unique fingerprints of ancient retroviral infection demonstrate relatedness if found in the genome of two different organisms.
1. Background
Viral infections. We all know and hate them. From the common cold to the common cold sore, they're annoying little buggers. The virus particle invades a cell, injects its proprietary genome, hijacks the cellular machinery and turns it into a virus-making factory. Upon successful completion of this process, the cell begins releasing fresh copies of the virus, often by bursting and dying.
Occasionally, however, something goes wrong in the integration process. When this occurs, the cell can survive, but selected portions of the viral DNA remain in its genome. These vary from infection to infection, not just from virus to virus.
In effect, the cell becomes permanently scarred and carries the unique, identifiable fingerprint of its assailant. (Source)
Since the markers are now in the genome, they become hereditary--hence, any descendant of the afflicted cell will contain the same exact viral fingerprint(s) at the same exact spot in its DNA.
Viral infections are, of course, not limited to the mucous membranes, blood stream and lungs. Under certain conditions, they can also infect a germ line (egg or sperm) cell. The odds of this cell now surviving to adulthood are astronomically low; not only does it have to survive a viral infection, but be one of the extremely lucky few that ever make it to merge with their counterpart and form a viable zygote.
However, this does occur. When it does, the resulting offspring has the unique viral fingerprint embedded in every cell in its body, what is known as an "endogenous retroviral insertion", henceforth referred to as an ERV. (Source)
ERVs can be easily identified by their extreme similarity to known retroviruses. They even contain vestiges of gag, env and pol genes that code for viral surface proteins (explanation of GEP protein coats, information on their presence in ERVs).
We've directly observed creation of new, identifiable ERVs by viral integration into cells in vitro, as well, so this process is as well-documented (if not more) as fossil formation. When it occurs in ordinary cells, it can cripple vital genes, subsequently causing cancer. High-risk papillomaviruses have been indicted as one of the leading causes of cervical tumors, for example. (Source)
Like any genes, the viral elements can then piggyback on the success of an individual and become established in the entire species. In small populations, this can easily occur via genetic drift (random establishment of selectively neutral or deleterious alleles). In larger ones, natural selection can work to establish the few ERVs that yield a benefit to their host.
Each step in the process is not very common, and has unique aspects that are extremely unlikely to be reproduced in two independent cases of ERV creation. Hence, when they all occur, the organisms possessing the ERV are marked with a molecular fossil that will prove indispensible in tracing their origin.
2. Evolutionary Implications of ERVs
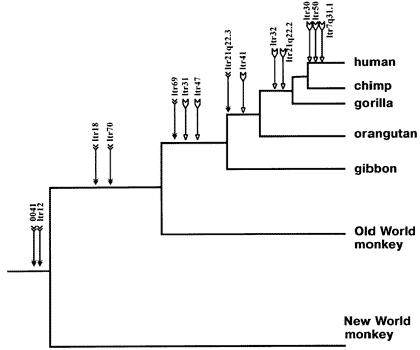
In a nutshell, this can serve as a "paternity test" of sorts: if you find the same endogenous retroviral insertion in two different organisms, the only viable explanation is that they inherited same from a common ancestor that, itself, possessed it. Offshoot species will have the same unlikely and easily identifiable ERVs, enabling us to construct accurate phylogenies from an independent line of evidence.
Human endogenous retroviral insertions in identical chromosomal locations in various primates. Notice just how well the standard evolutionary phylogeny, derived from fossil and anatomical evidence (humans and chimps closest, then orangutans and gorillas, then gibbons, then old world monkeys, then new world monkeys) is corroborated by this uniquely independent line of evidence.(Source)
As a side note, ERVs have also been used to reconstruct the relationships between dogs, jackals, wolves and foxes, among other species of domestic animal.
Needless to say, this offers a quick and easy way to experimentally test if any two similar organisms are unrelated. If they are, they won't share any ERVs. If you find the same one at the same place in their genomes, however, the hypothesis they originated independently is effectively falsified.
Due to the nature of the branching tree of life, ERVs shared between distantly-related organisms will have made their initial appearance in their common ancestor. Hence, they would have also been present as further, closer organisms split off from the lineage; if you find an ERV common to humans and monkeys, the ancestor of chimps must have had the insertion as well. Similarly, ERVs found in New World Monkeys and chimps will usually be present in humans. ERVs found in humans and gorillas should be found in chimps. And so on. This provides an additional way to test evolutionary predictions.
It should be noted that there are rare but observable genetic phenomena that can remove ERVs from the genome. However, we can almost always tell when this has occured, such as identifying the characteristic fingerprint of a deletion mutation right before the ERV 'should have' been found.
Nevertheless, the more basic question of 'are there shared ERVs between these species at all?' is a much more stringent, rock-solid test of evolution than the specific phylogenetic relationships individual insertions indicate, which can be erroneous on occasion.
When analyzed as a statistical whole, ERV insertion patterns will also recapitulate phylogenies drawn from independent data like the fossil record and comparative anatomy. This is a remarkable confirmation, and totally at odds with the predictions made by the hypothesis of independent origin of species.
As you can see by now, this is the strongest support for evolution I've ever come across; a truly powerful and damning smoking gun
Subscribe to:
Posts (Atom)
